

En el lliurament anterior vem veure com, des dels estudis de Ernst Haeckel, se sap que el desenvolupament embrionari recapitula la història evolutiva dels animals i ens va servir per presentar un bitxo d’actualitat (l’amfioxus). Anem a veure com relacionem tot això amb la obra més poderosa de Sauron: l’anell únic.
Entre d’altres poders l’Anell Únic “desenvoluciona” al seu portador. Mireu sinó al pobre Sméagol.
L’anell únic és capaç d’activar els mecanismes que tenen lloc durant el nostre desenvolupamnet... però en ordre invers! El seu portador comença a adquirir trets juvenils que es traduexien al principi en un augment de la longevitat i una aparença més juvenil que per edat pertocaria (en Bilbo n’és un exemple).
Però després la cosa va a més: els ulls van fent-se més grans respecte a la mida del crani i apareixen membranes digitals i, fins i tot membranes nictitants (la doble parpella de certs peixos, amfibis i rèptils). D’aquesta desenvolució n’és una prova (vivent?) en Gol•lum. I això ens porta a preguntar-nos el següent:
Fins on hagués arribat la desenvolució d’en Gol•lum sinó li haguessin pres l’anell? Hagués desenvolupat brànquies? On duria l’anell quan es trobés en fase d’amfioxus? Algun peix havia estat abans portador de l’anell? Si un peix es menjés l’anell, arribaria fins a la base simbiòtica dels eucariotes? Preguntes frikis. Preguntes sense resposta.
No perdem de vista l’amfioxus perquè ens permet enllaçar amb el tema de la següent setmana, la pedomorfosis.
29/6/08
Biotolkien II. Desenvolupament. L'anell únic.
24/6/08
Expedició a l'àrtic
Un amic del blog, Toni Pou, ha iniciat la seva presència en una expedició a l'artic. Des d'aquí li dessitjem tota la sort i que s'ho passi teta! Podeu seguir els seus passos aquí
Biotolkien II. Desenvolupament. Evo-Devo
L’estudi del desenvolupament embrionari ha deparat més d’una sorpresa. La universalitat dels gens que controlen la segmentació del cos de tots els vertebrats és un fet sorprenent que ens ha servit d’excusa per especular sobre l’origen evolutiu dels dracs (els famosos gens Hox, recordeu?). Però molt abans, el desenvolupament embrionari ja havia ofert una altra sorpresa: les etapes de la formació de l’embrió recapitulen la història evolutica de l’espècie a la que pertany. Aquest fet, estudiat i descrit pel gran biòleg (i millor dibuixant) Erns Haeckel, va sevir per apuntalar més la teoria de l’evolució.
Què vol dir “recapitular” l’evolució?
Com es pot veure a la figura, tots els embrions dels vertebrats comencen assemblant-se als embrions dels peixos. Després cadascun segueix el seu camí. Si agafem el cas d’un pollastre, aquest comença com un pexi, després passa a ser més com un amfibi i, després de superar la fase de rèptil, arriba al seu exclusiu camí d’au. Als humans, com a vertebrats que som, ens passa exactament el mateix.
Aquesta relació evolució-desenvolupament (Evo-Devo), comporta uns fets “estranys” en les nostres fases primerenques. Hi ha moments en què els nostres ulls, en proporció amb el crani, són molt més gran que a la fase adulta; a més, presentem membranes entre els dits de les nostres extremitats; membranes que acaben desapareixent (pel mecanisme de suïcidi cel•lular o apoptosi, ja tractat en aquest blog).
De fet, en uns primers estadis ens assemblem a uns rudimentaris amfioxus, uns estranys animalons que deriven d’un ésser que es va situar a la base evolutiva dels cordats (dins els cordats trobem els vertebrats). Aquests animals s’han fet famosos recentment perquè s’ha descodificat el seu genoma. Els diaris locals li han donat rellevància perquè ha estat portada del Nature, col•laborant-hi dos investigadors de la U.B. (també és cert que l’article el signen una trentena d’autors més, però un Nature és un Nature). Tindran gens Hox els amfioxus? Segur que sí.
Ups. M’he allargat massa. La setmana vinent veurem com l’anell únic es val de l’Evo-Devo per dur a terme algunes de les seves “accions”.
14/6/08
Biotolkien II. Desenvolupament. Teoria evolutiva dels dracs
La setmana anterior vam veure com els dracs tenien sis potes i com aquestes podrien haver sorgit de la mutació dels gens hox. Evolutivament això generava tot un seguit de problemes que requerien d’una teoria per a poder explicar-los. I aquí us en va una.
1) L’ancestre directe dels dracs ha de ser un rèptil de mida petita, com un llangardaix o un pèl més gran, segurament verinòs (per justificar el verí que semblen tenir tots els seus descendents, i fins i tot, la possibilitat de generar foc).
2) Algun d’aquests ancestres va patir una mutació en els seus gens hox que va fer que es dupliquessin les extremitats davanteres. Aquest rèptil hauria de ser bíped per a que les quatre potes davanteres no li supossessin cap molèstia a l’hora de desplaçar-se.
3) Aquesta mutació es va mantenir i va anar derivant a extremitats més especialitzades. Mentres les més davanteres continuaven sent potes, que podríen fins i tot permetre’ls-hi agafar coses i trepar, les del mig van anar desenvolupant una membrana digital. Potser al principi servia per perdre més ràpidament la calor o per guanyar-la si vivien en zones assoleiades (recordem que els rèptils solen ser de sang freda, al menys els actuals).
4) Aquest llangardaix de sis potes trepador començaria a passar d’arbre a arbre gràcies a les membranes del seu tercer parell d’extremitats.
5) Quan la mida encara no era tan bestial van acabar desenvolupant tots els mecanismes necessaris per al vol.
6) Més tard la seva mida augmentà donant lloc als primers dracs pròpiament dits. Aquest ja deurien ser de sang calenta per poder dormir dins de fredes coves.
7) D’aquests dracs derivarien tota la resta, els quals anirien perdent extremitats.
És tan sols una teoria difícilment demostrable... algú té alguna altra?
La setmana vinent parlarem d’embriologia i de com els embrions ens aporten informació de la història evolutiva de l’espècie. També veurem com l’anell únic va poder alterar la fisionomia d’en Gol•lum d’aquesta manera.
10/6/08
Cristina Garmendia
Dissabte, a El País, va aparèixer publicada una entrevista de Berna González Harbour a la nova ministra, Critina Garmendia, del nou ministeri de Ciència i Innovació. Si la creació del ministeri ja va ser tota una declaració d'intencions del nou executiu, posar al capdavant una biòloga amb doctorat, anys en la investigació pública i que havia donat el salt a la privada, va ser un reconeixement explícit a la situació de la "carrera" científica en aquest país.
L'entrevista em va agradar. La ministra es mostra directa i amb les idees clares, sense refusar parlar dels sous ridículs que perceben els investigadors i docents universitaris d'aquest país (encara que, de fet, falta una referència clara i directa que digui que és un dels principals problemes perquè els cervells fugits tornin). És també ambiciosa, proposant un canvi en les inversions d'aquest país, del maó a la probeta. Veurem si no peca de massa idealisme.
A l'estrevista s'incideix en la necessitat de posar alguna universitat espanyola entre les 100 primeres del món. Segur que han pensat -o estan pensant molt en això- però un simplíssim pas a considerar seria aconseguir que els doctorats que surten de les nostres Universitats no ho facin tots amb la màxima puntuació. Desmereix a la pròpia Universitat. Si no hi ha pluralitat en les notes, tots els doctors "cum laude" d'aquest país no són igual de brillants, són igual de mediocres. Realment desconec la situació en altres països, però espero que no sigui com la que es dóna aquí, on una Universitat veu com un segell d'Excel•lència que tots els doctorats siguin cum laudes per unanimitat, sense importar-los que una lectura superficial de 5 o 6 de les seves tesis reveli diferències notabilísimas entre elles. Però, clar, qui s'atreveix a puntuar a la baixa el treball d'un "alumne" d'un cap de grup el qual pot puntuar dintre de mig any el treball del teu "alumne"? Com podem puntuar el treball de quatre, perdó, cinc o sis o més anys? Algunes preguntes és bo plantejar-se-les de vegades. D’altres és millor no plantejar-se-les mai.
Un altre tema absent a l'entrevista va ser la necessitat d'un augment en la cultura científica d'aquest país. La nova assignatura de Ciències per al Món Contemporani (que, increïblement ha irritat a sectors anquilosats i reaccionaris d'aquest país que preferirien Ciències per al Món de l’Edat Mitjana) pretén començar a omplir aquest buit, però una major presència de la Ciència, ben explicada, als mitjans de comunicació, així com activitats culturals i tallers promoguts pels governs, haurien de ser un pilotis en aquest "canvi" que es proposa des del ministeri. Si la ciutadania entén què s'està fent, considerarà un avanç la Ciència; si no s'informa correctament, qualsevol persona ignorant o malintencionada podrà tergiversar les investigacions de científics espanyols per a cometre actes atroços contra el seu treball amb la connivència d'un poble desimformat. Cosa que ja ha passat.
En fi, per demanar que no quedi.
8/6/08
Biotolkien II. Desenvolupament. Dracs
En el lliurament anterior vam presentar els gens Hox, implicats en el desenvolupament embrionari dels animals. Aquests gens determinen què farà cadascun dels segments del cos de l’animal en formació: on aniran les ales, les potes, els braços, etc. Mutacions en aquests gens donen lloc a alteracions espectaculars, com les que vam veure en la mosca del vinagre. I d’aquí anàvem a passar als dracs.
No sé si us ho havieu plantejat, però els dracs, a diferència de la resta de rèptils, tenen sis extremitats. Un possible origen del segon parell d’extremitats superiors el trobaríem en una mutació en algun o alguns gens hox, aquells que determinen la generació de “braços” en un lloc determinat. Duplicant aquesta ordre obtindríem dos segments d’on sortirien parells de “braços”. Dos d’aquests braços haurien patit una selecció evolutiva que els hagués dut a desenvolupar una membrana interdigital que acabaria transformant-se en ales. Però aquí sorgeixen moltes incògnites:
No desesperem. De tot es pot treure una teoria... i la setmana vinent us en plantejaré una.
2/6/08
Lab Basics (4): Fotocopiant àcids nucleics (II)
L'entrada anterior de la sèrie va descriure una de les tècniques mestres en els laboratoris de biologia mol·lecular (i d'altres) de tot el món: la PCR. Convé tenir bé clar el concepte perquè anem a comentar un parell de coses més al respecte.
Per exemple, ens vam centrar només en explicar com amplificar DNA. Què ocorre quan partim de mol·lècules de RNA purificat, que són de cadena senzilla? El procés en cadena descrit anteriorment era vàlid per a les mol·lècules de doble cadena. No hi ha problema: tornem a aplicar enzims disponibles a la natura per a solucionar-ho, en aquest cas la transcriptasa inversa (o retrotranscriptasa) vírica. No és més que una polimerasa particular que utilitza RNA com a motlle. El procediment per a amplificar RNA més comú és el següent: posem en el mateix tub certa quantitat de RNA purificat juntament amb seqüències curtes de moltes timines (poliT); aquests oligonucletids (...TTTTT...) s'hibridèn a alta temperatura (per a això utilitzem també un termociclador) amb la cueta d'adenines (---AAAA---) que tenen les mol·lècules de RNA sintetitzades en el nucli (cueta que les permet sortir al citoplasma per a la transcripció sense que les degradin). El paper de la poliT és anàleg al dels cebadors per a amplificar ambdues cadenes del DNA. Després de la hibridació amb les poliT, h'hia ha prou d'afegir a la reacció la trancriptasa inversa (més cara que la convencional) amb el seu respectiu buffer o tampó, nucleòtids i completar el volum amb aigua. Es deixa actuar la transcriptasa inversa (o reversa) a durant una hora a la temperatura adequada i s'obtenen com a resultat grans quantitats de molècules de cadena senzilla, equivalents al RNA inicial, però amb timina en lloc d'uracil. És a dir, similar al DNA de tota la vida però sense introns (seqüències no codificants) i de cadena senzilla. Aquesta molècula creada ex professo en laboratori es coneix com cDNA (DNA còpia), i és una manera immillorable d'obtenir una seqüència codificant de DNA (gen) sense presència d'introns, que es pot purificar i usar per a mil tècniques posteriors més (algunes de les quals explicarem d'aquí a poc). El cDNA obtingut pot servir al seu torn de base per a una amplificació convencional de DNA: en la primera reacció com és obvi només es pot amplificar la cadena que existeix, però la cadena senzilla vella formarà una doble cadena amb la nova i en les reaccions següents partim de la situació normal: molècules de doble cadena llestes per a amplificar.
Aquesta reacció descrita es diu RT-PCR (PCR de transcriptasa reversa), no confondre amb la PCR a temps real (Real Time PCR), sistema que serveix per a quantificar àcids nucleics (DNA i RNA). És una tècnica més complexa que requereix un termociclador especial, capaç de mesurar fluorescència. La idea és intercalar en la cadena de l'àcid nucleic una molècula que doni senyal fluorescent (fluoròfor), que es pugui detectar i quantificar i el senyal del qual sigui proporcional a la quantitat de producte de la PCR. Existeixen variacions i complicacions en la tècnica però el més habitual és detectar específicament una seqüència utilitzant oligonucleòtids marcats amb fluoròfors (sonda), que contenen el fluoròfor en un extrem i una mol·lècula segrestadora de la fluorescència del fluoròfor en l'altre (quencher). El contingut de la reacció de PCR és similar a l'habitual, també existeixen cebadors d'inici de l'amplificació. La sonda és complementària a un fragment de la seqüència diana, però la polimerasa amplifica a partir del cebador.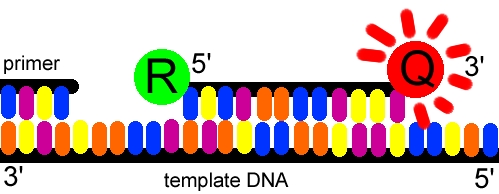
Quan la polimerasa es topi amb la sonda, separarà el fluoròfor de la resta de la sonda i es podrà detectar la seva fluorèscencia (lliure del quencher segrestador), que augmentarà exponencialment segons el nombre de còpies del DNA motlle.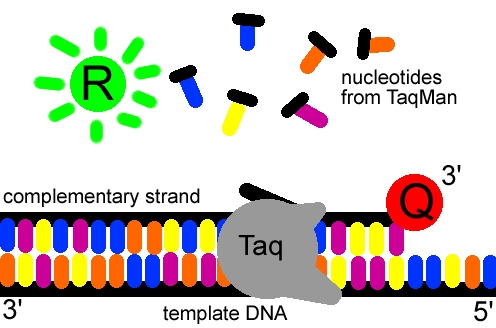
Es poden usar vàries sondes amb fluoròfors diferents alhora. Per a la quantificació s'analitza la corba d'amplificació, que consta de 3 fases: una en la qual el producte encara no es pot detectar, la fase d'acumulació exponencial del producte i la fase de saturació de la reacció. El nombre de cicles necessaris perquè es produeixi un augment de fluorescència significatiu pel que fa al senyal basal (cicle llindar) és inversamente proporcional a la quantitat inicial de mol·lècules motlle. Mitjançant els càlculs pertinents (ja seria molt llarg explicar-ho) es pot obtenir una quantificació precisa del nombre de mol·lècules inicials en la reacció.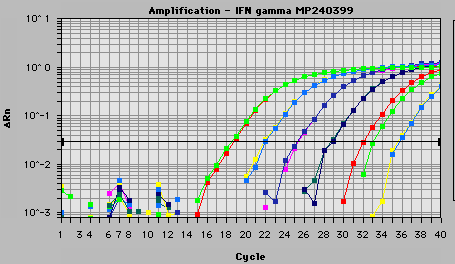
Us podeu fer idea de la sofisticació creixent d'aquest tipus de tècniques. Hi ha mil i una variants de la PCR, però no us torturo més.
1/6/08
Biotolkien II. Desenvolupament. Els gens Hox

En aquesta nova entrega de Biotolkien ens centrarem en el desenvolupament embrionari i parlarem de dracs, hòbbits, huargos, mûmakils, i d’en Gol•lum. Veurem com poden haver sorgit les ales dels dracs, com l’anell pot haver influït en la forma tan estranya d’en Sméagol i com alterant els ritmes de desenvolupament es poden aconseguir individus i espècies diferents de les seves espècies ancestrals.
Si em permeteu començarem pels dracs i les seves ales.
De dracs en el món d’en Tolkien n’hi ha de molts tipus: en forma de cuc, amb braços i cames, de gel, de foc, de vent. Que volen, que no... però el propi Tolkien ens deixa ben clar que els més perillosos són els dracs alats de foc. No entraré en el mecanisme biològic que pot conduir a treure foc pels queixals (o al menys no en aquesta entrega de Biotolkien), però sí en el mecanisme pel qual poden haver sorgit les ales d’aquests animals. I la solució l’hem de buscar en les mosques.
Com molts de vosaltres sabeu la mosca del vinagre és un organisme model que s’utilitza en genètica des de fa molts i molts anys. Aquestes mosques es reprodueixen amb relativa rapidesa produint gran quantitat de descendents, el que la fa ideal per veure com es transmeten els caràcters durant diferents generacions. A més, és un organisme en què, de manera natural, o induïts, trobem una quantitat impressionant de mutacions diferents. Una d’aquestes mutacions és la bithorax. Les mosques normals tenen un parell d’ales grans i un parell d’ales més petites en el següent segment del cos que s’anomenen halteris. Les bithorax tenen quate parells d’ales. Dos al segment que els hi pertoca i dos enlloc dels halteris. Aquest canvi tan radical és degut a que durant el desenvolupament embrionari de la mosca es van definint els diferents segments que formaran el seu cos. Així va quedant clar on hi haurà els ulls, on la boca, les potes, les ales, els halteris, etc. Si en algun moment hi ha un error poden sortir coses on no tocarien. Per exemple, les mosques antennapedia tenen potes en comptes d’antenes per una mutació en un gen de la familia Hox. Aquesta familia de gens controla la segmentació dels cossos en els animals. 
I quan dic animals també incloc els vertebrats. Els gens de la familia Hox dels vertebrats (molt semblants als de mosques) són els que determinen la longitud del coll de cada espècie animal (controlant quants segments “cervicals” tindran), on van els braços, les cames, i moltes altres coses. I aquí arribem als dracs. Però ho haurem de deixar per al proper lliurament.
23/5/08
granotamandra
Els amfibis que coneixem avui en dia es classifiquen en tres grups: granotes, salamandres i cecílies. El que encara no està clar és com aquests tres grups han evolucionat, sobretot perquè no s'han trobat gaires fòssils que ens ho expliquin. Fins ara, l'única manera que hi havia d'estimar quan es van separar les granotes i les salamandres era guiant-nos pel temps que ens marca el rellotge molecular, un mètode que permet calcular el temps que fa que dues espècies han divergit del seu avantpassat comú basant-se en el número de diferències genètiques entre elles. El rellotge molecular situava aquesta separació entre fa 300 i 350 milions d'anys.

Ara, es publica a Nature la troballa d'un fòssil que omple el buit que hi havia entre els amfibis més primitius i els primers membres de la família de granotes i salamandres, ja que comparteix algunes característiques pròpies de cada un dels dos grups. El crani, per exemple, és semblant al de les primeres granotes, mentre que els ossos del "peu" que s'han trobat són característics de les salamandres. La columna és un forma de transició, és a dir, té un número de vèrtebres que es troba entre el dels amfibis més primitius i les primeres granotes i salamandres.
Totes les característiques observades en aquest fòssil, el Gerobatrachus hottoni, han permès situar el moment de la divergència entre aquests dos grups fa entre 270 i 280 milions d'anys. És a dir, que la separació entre granotes i salamandres és més recent que no ens pensàvem. A més a més, l'article estima que la separació entre el grup format per granotes i salamandres i el grup de les cecílies va tenir lloc fa, com a mínim, 328–335 milions d'anys.
11/5/08
Lab Basics (3): Fotocopiant àcids nucleics (I)
A l'entrada anterior de la sèrie vam veure com obtenir DNA (i RNA) a partir de material biològic (cèl·lules en cultiu, sang). Ara que tenim les nostres cadenes de DNA resuspeses en un tubet amb líquid, què és el que podem fer amb elles?
Lamentablement poc, perquè aquest material que hem obtingut és el material genòmic complet de l'individu (poc pràctic per a treballar), i a més està poc concentrat. El més corrent és que a nosaltres ens interessi treballar amb una part de la seqüència d'aquest genoma, ja sigui en el gen o gens implicats en la nostra investigació o en una altra regió concreta no codificant.
Aquesta pretensió va poder convertir-se en realitat a les acaballes dels 70 quan, aplicant en laboratori els mecanismes de replicació del DNA inspirats en els que tenen lloc en la cèl·lula, es va dissenyar la tècnica de la reacció en cadena de la polimerasa (PCR) per a obtenir ingents quantitats del fragment de seqüència desitjat.
En la naturalesa i en el laboratori es parteix del mateix, posem de moment, ADN nuclear. El primer que en la cèl·lula ocorre per a replicar aquest material és separar ambdues cadenes de la molècula d'ADN, mitjançant enzimes helicases. Però separar els ponts d'hidrògen entre les bases nitrogenades de la cadena de DNA també es pot fer a altes temperatures, entre 90-98ºC (a més ponts d'H que calgui trencar, més temperatura). Això es diu desnaturalització.
Una vegada amb les cadenes separades, la síntesi de la nova molècula, còpia de l'original, es produeix en sentit 5'-3' de la cadena, i partint d'un fragment previ de cadena complementària a l'original que proporciona l'extrem -OH 3' lliure per a anar afegint nucleòtids. Aquests fragments en la cèl·lula els crea la primasa, i la síntesi de la cadena copia,l'enzima polimerasa.
Aquests petits oligonucleòtids inicials de cadena senzilla (encebadors, o primers) es creen en laboratori i es poden dissenyar a mà o amb programaris disponibles en internet i demanar a cases especialitzades (uns 12 euros cada oligonucleòtid de 20 parells de bases, que és el més usual, així que cada parella de complementaris et surt normalment per menys de 25 euros). Es dissenyen perquè siguin complementaris a un fragment de la regió d'interès i serveixin de base per a amplificar una i una altra cadena de la molècula motlle en ambdós sentits (en l'esquema de baix ho entendreu millor). Es lliuren liofil·litzats i es preparen per a utilitzar-los a una concentració final de 10-20 picomols per microlitre de reacció.
Per a unir-los a la cadena que vulguem copiar utilitzem també la temperatura: segons la seqüència de cada oligo aquesta temperatura d'hibridació varia entre 45-64 graus aproximadament(entre 50-58ºC és el més habitual). Com aquesta unió a les cadenes motlle només es produeix si ambdues seqüències són pràcticament idèntiques, ens assegurem que només copiarem la regió que nosaltres hem delimitat mitjançant el disseny dels nostres encebadors.
La síntesi de la cadena motlle la hi deixem a polimerases obtingudes de bacteris extremòfils, altament resistents i actius a les temperatures a les quals estem maltractant el DNA, i que amplifiquen la cadena normalment a 68-74ºC, segons la polimerasa. Una unitat d'enzim és aquella quantitat que converteix 1 mol de substrat per segon. La polimerasa es ven en tubs de 500 a 10000 unitats (500 unitats et poden costar entre 65-120 euros) i s'empra una concentració final de 1 unitat per microlitre de reacció.
Aquests enzims necessiten per al seu funcionament ions bivalents (magnesi normalment), que s'aporta en forma de clorur agregat o no a un tampó o buffer que manté un pH adequat per a la reacció. El tampó i el magnesi se sol vendre en kits juntament amb la polimerasa corresponent. I per suposat, es necessiten nucleòtids (desoxinucleòtids trifosfat o dNTPs), els maons necessaris perquè la polimerasa pugui anar construint la nova cadena còpia.
Així doncs necessitem per a la reacció una quantitat suficient de DNA còpia (50-100 ng és el més usual), que barrejarem en un tub en condicions d'esterilitat per a evitar barreges amb material genètic forà amb la parella d'oligonucleòtids, la polimerasa, el seu tampó i magnesi, nucleòtids i aigua destil·lada estèril i sense nucleases fins a un volum final de 25-100 microlitres.
Fixeu-vos que en un cicle d'amplificació que hem descrit (desnaturalització, hibridació, síntesi) per cada molècula motlle s'obtenen dues molècules noves (cadascuna amb una cadena motlle i una altra còpia). Mitjançant cicles successius d'aquesta reacció podem obtenir 2 elevat a n molècules de DNA del fragment que a nosaltres ens hagi interessat, on n és el nombre de cicles (30-40 és el més normal; més enllà la quantitat de DNA produït no és exponencial per esgotament dels productes de la reacció). I amb aquestes quantitats de DNA obtingut sí es pot treballar.
Antany tots aquests canvis de temperatura necessaris per als cicles d'amplificació els realitzava un operari canviant els tubs d'unes cubetas calentes a unes altres. Afortunadament de seguida es van emprar termocicladors, una espècie d'estufes amb temporitzador en les quals es pot introduir els tubs i programar el temps que vulguem que el termociclador escalfi o refredi per a cada fase del cicle. Un termociclador modern bàsic ronda els 6000 euros, encara que els hi ha molt més sofisticats.
Per a la desnaturalització se solen emprar pocs segons (mig minut o així); per a la hibridació, 30-40 segons haurien de ser suficients; el temps de síntesi depèn de la longitud que nosaltres vulguem amplificar (delimitada pels oligonucleòtids dissenyats): normalment una polimerasa típica amplifica 1000 parells de bases en un minut; se solen afegir alguns minuts extres, fora del cicle, perquè les cadenes deixades a mitjes s'acabin de sintetitzar, i finalment la reacció es para deixant els tubs a 4 graus.
O sigui, que en una hora i dos quarts o dues hores obtens 2 elevat a 35-40 molècules del teu tros de DNA desitjat en un tub (excepte la gent de la sèrie CSI, que ho obté en 1 minut escàs per exigències del guió).
La setmana que ve complementarem aquesta informació amb alguns altres detalls sobre la PCR i el que es fa després amb els tubs de reacció acabada.







{kind=link}
{kind=link}
{kind=link}